雌雄异花甜瓜经AgNO3诱导后的雄蕊发育和差异基因表达 
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2011 年, 第 9 卷, 第 10 篇 doi: 10.5376/mpb.cn.2011.09.0010
收稿日期: 2010年09月17日 接受日期: 2010年12月28日 发表日期: 2011年01月31日
李晓明等, 2011, 雌雄异花甜瓜经AgNO3诱导后的雄蕊发育和差异基因表达, 分子植物育种 Vol.9 No.10 (doi: 10.5376/mpb.cn.2011.09.0010)
本研究利用cDNA-AFLP技术对AgNO3诱导后的甜瓜雌雄异花系'RH107'mRNA进行指纹图谱分析,320对引物组合共分离到差异表达的TDF (transcription derived fragment, 转录衍生片段) 131个。对其中的28个TDF进行了克隆、测序和序列分析。研究结果表明,在获得的16个有效序列中,6个与GenBank中功能已知基因同源性在72%和100%之间,其功能涉及信号传导、转录调控和能量代谢等;5个与功能未知蛋白的核苷酸序列或氨基酸序列有同源性;2个与已知表达序列标签(EST)有同源性,但功能未知;3个未找到同源序列。由此推断,Ag+诱导表达差异片段可能分别参与了信号转导、基因表达调控和能量代谢等过程,从而参与调节花器官性别分化。
瓜类作物的性型表现多样,遗传机制复杂,易受环境条件影响,一直是科研和生产上关注的热点。Owens 等(Owens et al., 1980)创制甜瓜新种质——全雌系WI998时,采用硝酸银处理幼苗诱雄,用以解决自交留种问题。此后,Ag+成功诱导黄瓜、苦瓜、西葫芦等作物雌性系雄化(Ando et al., 2001; 杨等, 2004; 戚艺军等, 2004),逐渐成为一项亲本繁殖和种子生产的成熟技术。甜瓜ACC合成酶基因CmACS-7的克隆,揭示了该基因一处活跃位点上的C-T突变改变了氨基酸序列,从而决定了雄蕊的发育与否(Boualem et al., 2008)。Ag+处理能够诱导雄蕊发育,达到雌性系自交留种的目的,但是雌性系的后代依然保持了雌性系特征,说明Ag+处理没有引起CmACS-7基因的突变,却改变了雄蕊发育的最终结果。因此推测,Ag+必然是作用在了多基因协同控制雄蕊发育的网络系统中的某一个或几个关键点上,但是到底发生在性别决定基因的上游还是下游,以及如何对雄蕊发育过程进行影响的,目前这些机制尚不清楚。
近年差异表达基因研究方法不断丰富和完善,Northern杂交、实时定量RT-PCR、表达序列标签(Expressed sequence tags, ESTs)、mRNA差异显示技术、抑制差减杂交(Suppression subtractive hybridization, SSH)、基因芯片(Gene chip)等等,使得人们能够从转录水平对差异表达基因进行分离和功能鉴定。cDNA-AFLP是以AFLP为基础的基于目的片段长度的RNA指纹分析技术,合适的酶切组合几乎可以检测所有表达的基因,具有重复性好、假阳性率低等特点(Bachem et al., 1996),已被广泛用于差异表达基因的分离和鉴定、基因表达特性等方面的研究,在分离芥菜(Fusco et al., 2005)、大麦(Leymarie et al., 2007)、番茄(Frank et al., 2009)等作物发育和逆境响应相关基因方面发挥了巨大作用。
本部分试验以AgNO3处理雌雄异花甜瓜生长点,诱导甜瓜雌花雄蕊发育,利用cDNA-AFLP基因差异表达技术分离与性别分化相关的特异表达转录衍生片段,为进一步克隆相关基因、揭示甜瓜性别分化分子机理奠定基础。
1结果与分析
1.1 AgNO3处理对甜瓜性型的影响
雌雄异花试材‘RH107’经200 mg•L-1 AgNO3处理后,对性别表达产生影响,表现为“促雄”作用,即在基部的1-4朵雌花中生长了3-5枚雄蕊,性别由雌花转变成两性花(图1)。AgNO3处理对雄花数目和节位无显著影响。
(1).png) 图1 雌花经AgNO3诱雄后雄蕊发育 Figure 1 Morphology of the developed stamens in a female flower reduced with AgNO3 |
1.2 cDNA-AFLP分析
320 个引物组合进行选择性扩增,在处理池和对照池间所扩增出多态性片段为131个,占TDF总数的2.2%,每对引物平均扩增出差异TDF为0.40个。在差异TDF中增强表达21个,占总数的16.0%;特异表达37个(图2),占总数的28.2%。另外73个TDF在AgNO3诱导下表达被抑制,占总数的55.7%(图3)。
 图2 部分上调表达的片段 Figure 2 Up-regulated fragments by cDNA-AFLP analysis |
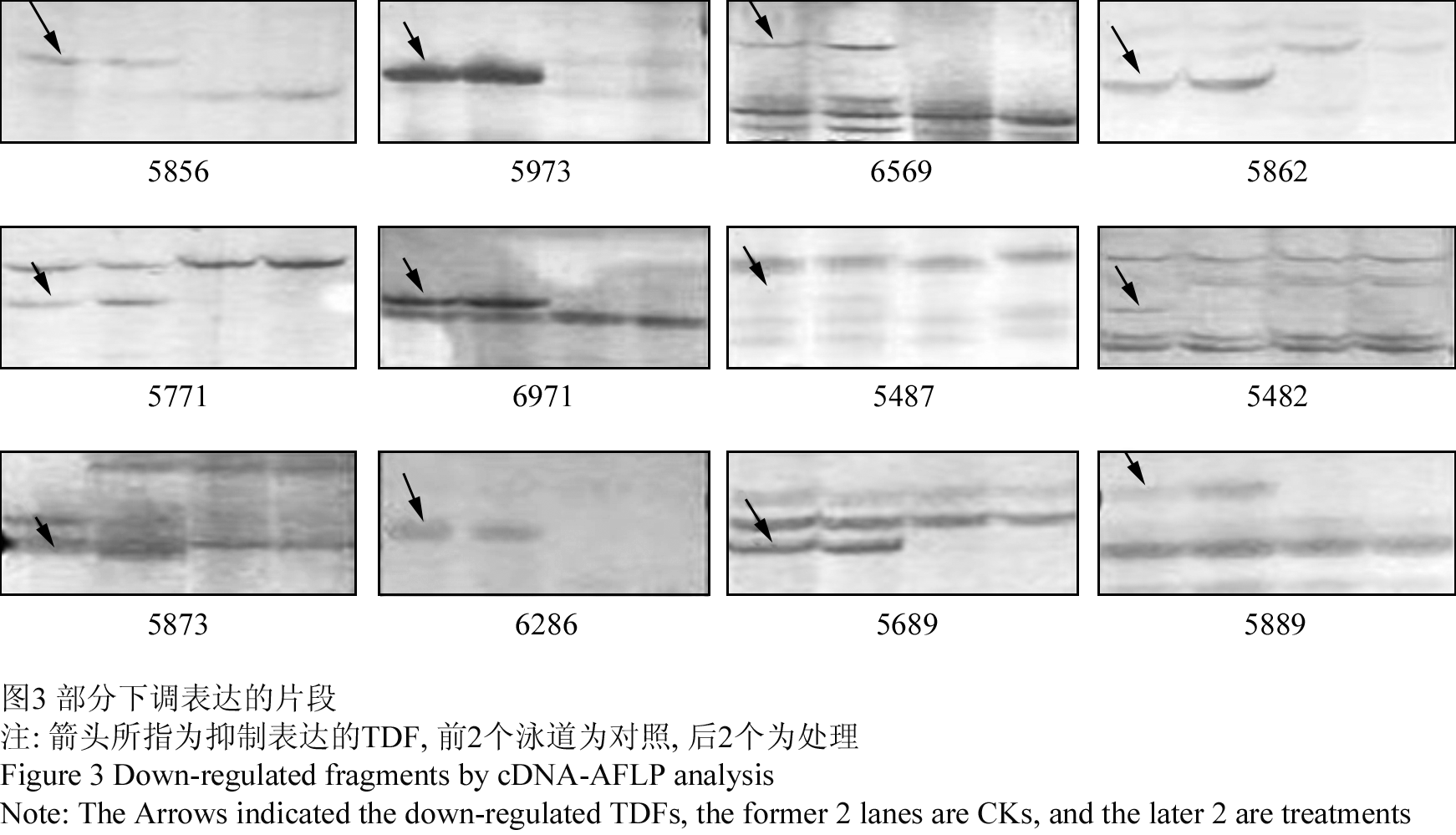 图3 部分下调表达的片段 Figure 3 Down-regulated fragments by cDNA-AFLP analysis |
1.3差异表达片段克隆测序及序列分析
选取典型的特异表达和抑制表达TDF 28条进行回收和二次PCR扩增。多数引物组合产物分子量与预期结果相同,而且为单一条带。少数引物组合出现非特异性弱带,此时切取主带回收,通过T-A 连接到pGM-T载体,重组质粒转化大肠杆菌TOP10后,经菌体PCR扩增后挑选阳性测序,共得到16个有效序列。利用NCBI的BLAST工具与 dbEST数据库,进行特异表达基因同源性比较以及功能预测,结果显示,6个与已知生物学功能的片段具有较高同源性,其中分别涉及代谢(TDF2-12- 3, TDF3-11-6, TDF3-4-2)、信号传导以及细胞膜运动(TDF2-9-4, TDF3-9-1)、转录调控(TDF4-13-6),7个TDF具有较高同源性但缺少生物学功能,3个在NCBI中未找到同源序列(表1)。
 表1 差异表达TDF的功能预测 Table 1 The predictive function of the differential expression TDFs |
1.4 RT-PCR分析
为进一步验证cDNA-AFLP结果的可靠性,对部分差异表达的TDF进行了半定量RT-PCR验证。结果表明,TDF 2-12-3和TDF 3-9-1在Ag+作用下被诱导表达,RT-PCR和cDNA-AFLP结果吻合(图4),该方法所得结果可靠。
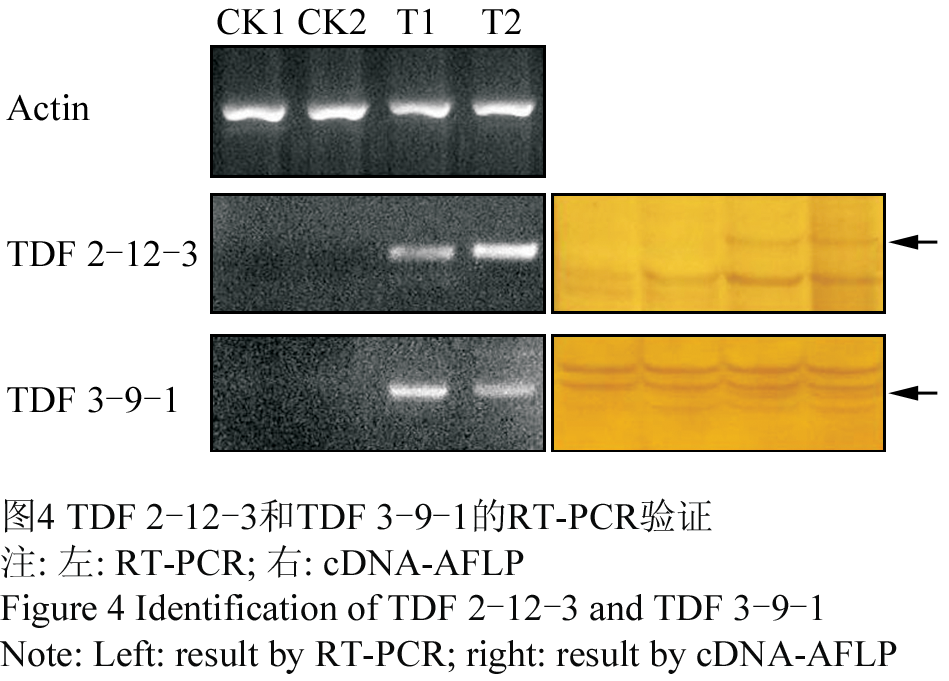 图4 TDF 2-12-3和TDF 3-9-1的RT-PCR验证 Figure 4 Identification of TDF 2-12-3 and TDF 3-9-1 |
2讨论
Beyer等(1976)认为Ag+是乙烯作用的竞争性抑制剂,能抑制乙烯的部分氧化代谢途径,进而抑制乙烯原初反应,最终抑制黄瓜雌花产生。汪俏梅、戚艺军等(1997; 2004)认为Ag+作为乙烯生物活性抑制剂而抑制乙烯活性,可能是通过竞争结合细胞上乙烯受体蛋白,起到削弱或降低乙烯作用的效果,从而诱导苦瓜性型雄化。对于拟南芥乙烯受体研究表明,位于内质网膜上的乙烯受体ETR1蛋白与乙烯结合必须依赖金属离子亚铜(Cu+)作为辅因子才能结合乙烯(Rodriguez et al., 1999),引起一系列乙烯信号感知反应。Ag+作为乙烯受体竞争性抑制剂,能够替代亚铜与乙烯结合,但是不能引起乙烯反应,从而阻断乙烯的信号转导(Schaller et al., 1995a, 1995b)。Theresa (2002)用AgNO3和Ag2S2O3分别处理雌雄异体植物白花蝇子草(Silene latifolia Poiret subsp. alba)的雌株,发现这种植物虽然对外源IAA、CTK、GA、ETH都不敏感,却能分别被两种Ag+制剂诱导后促进雄蕊的发育,在雌花上长出较长的花丝和较大的花药室,并完成减数分裂产生小孢子,只是花粉不能发育成熟。Theresa又用乙烯受体抑制剂1-MCP、乙烯生物合成抑制剂反式-环辛烯、AVG、CoCl2不同浓度做进一步的试验证明,这些制剂都能在白花蝇子草上发挥各自的抑制乙烯作用,但却无一能像Ag+那样产生诱导雄蕊的作用。因此得出结论:Ag+对于白花蝇子草的诱雄作用不能被解释为乙烯合成和信号传递受到抑制,暗示了这种效应可能存在另外的机制。本试验中采用Ag+作为甜瓜雌雄异花系的诱雄剂,客观上起到了诱雄作用,然而未能发现Ag+与乙烯相关基因表达或乙烯释放量之间的规律(数据未列出),但却发现一些与几类酶具有高度同源性的TDF被诱导而检测到差异表达,与Theresa的研究结果存在某种类似。
长期以来Ag+被看做许多带有巯基的酶的活性抑制剂(Snodgrass, 1960),能够抑制质膜和线粒体膜上ATPases的活性(Knee, 1992)。Ag+和巯基的密切关系对于酶活非常重要,这也说明植物的许多生理过程能够为Ag+所修饰改变,其中的某些过程可能涉及到雄蕊的发育(Theresa, 2002)。王日升等(2009)采用cDNA-AFLP对AgNO3诱导下纯雌系苦瓜花蕾发育的差异表达基因进行了分析,获得一个与植物性别分化有关的细胞色素P450家族基因有高度同源性的片段McCYP72A1。本文分离鉴定的差异表达cDNA片段与几类代谢酶基因的核苷酸序列或氨基酸序列存在一定程度同源性,推测Ag+处理后植物体很可能从非乙烯的其他途径对Ag+的诱导应答,进而引发了一系列信号传递网络式反应,“无意”间激活了某些雄蕊发育促进因子或者关闭了某些雄蕊发育抑制因子。但是,本文所获得的只是少部分差异片段而不是完整的cDNA,有待于通过RACE 等方法获得全长cDNA,对其表达特性和功能进行分析和鉴定,才有可能最终明确这些片段与雄蕊发育之间的确切关系。
3试材与方法
3.1试材及处理方法
雌雄异花品系‘RH107’,日光温室育苗。2~3叶1心时,用200 μg•L-1 AgNO3水溶液喷施全株。喷施后4、12、24、48和72 h分别取生长点,液氮速冻后保存于-80℃备用。以喷施蒸馏水为对照。各时间点设2次重复。植株开花后调查性型。
3.2 RNA提取及cDNA合成
RNA simple Total RNA Kit (TIANGEN公司, 离心柱型)进行总RNA提取,RQ DNase I (Promega公司)消化残留的基因组DNA。根据测得的RNA样品浓度,将RNA分别等量混合,构建2个对照池和2个处理池。M-MLV RTase cDNA Synthesis Kit (TaKaRa公司)用于双链cDNA的制备,步骤参照说明书。
3.3 cDNA-AFLP分析
使用EcoR I和Mse I两种限制性内切酶进行cDNA双酶切后连接接头,引物E00和M00用于预扩增,接头为EcoRⅠ和MseⅠAdaptor。选择性扩增引物Mse I 30个和EcoR I20个(各带3个选择性碱基),共计320个引物组合,用于选择性扩增,具体参照刘志勇等(2008)的方法。
3.4差异片段功能分析
挖带回收,加入PCR体系中,利用相应的引物组合扩增差异条带,进行二次PCR扩增。切取目标条带,AxyPrepTM DNA Gel Extraction Kit (Axygen公司) 回收特异片段,连接、转化,上海生工测序。对获得的序列结果,应用在线BLAST程序,在GenBank核酸序列数据库或EST数据库中搜寻同源序列,进行可能的功能分析。
3.5 RT-PCR分析
以内参基因CMACTIN作为对照,选取TDF 2-12-3和TDF 3-9-1作为RT-PCR验证的片段,设计合成特异性引物(表2),由Invitrogen生物工程公司合成。利用Superscript (Invitrogen)逆转录酶,以Oligo dT (18)为逆转录引发引物,逆转录合成cDNA第一链。
表2 用于RT-PCR的引物及序列 Table 2 Primers and their sequences used in semi-quantity RT-PCR |
作者贡献
李晓明是实验设计和实验研究的执行人,并主要负责论文写作;刘志勇是实验设计人,同时指导数据分析;冀瑞琴指导数据分析并提供实验引物;魏鹏参与论文写作;冯辉是项目的构思者及负责人,指导实验设计,数据分析,论文写作与修改。全体作者都阅读并同意最终的文本。
致谢
本研究由国家科技支撑计划(2006BAD01A07)、辽宁省教育厅博士访学计划(辽教发【2009】493号)资助。作者感谢两位匿名的同行评审人的评审建议和修改建议。本文中提到了我们实验中涉及的有关试剂供应商和测序服务商,这并非我们为这些试剂供应商和测序服务商的产品和服务提供推荐或背书。
参考文献
Ando S., Sato Y., Kamachi S., and Sakai S., 2001, Isolation of a MADS-box gene (ERAF17) and correlation of its expression with the induction of formation of female flowers by ethylene in cucumber plants (Cucumis sativus L.), Planta, 213(6): 943-952 doi:10.1007/s004250100571
Bachem C.W.B., Hoeven R.S., and Bruijn S.M., 1996, Visualization of differential gene expression using a novel method of RNA finger Printing based on AFLP: analysis of gene expression during potato tuber development, Plantjournal, 9: 745-753 doi:10.1046/j.1365-313X.1996.9050745.x
Beyer E., 1976, Silverion: a potent antiethylene agent in cucumber and tomato. HortScience, 11: 195-196
Boualem A., Fergany M., Fernandez R., Troadec C., Martin A., Morin H., Marie-Agnes S., Fabrice C., Jonathan M. F., Michel P., Michael D., Purugganan, Catherine D., Abdelhafid B., 2008, A Conserved mutation in an ethylene biosynthesis enzyme leads to andromonoecy in melons. Science, 321: 836-838 doi:10.1126/science.1159023
Frank G., Pressman E., Ophir R., Althan L., Shaked R., Freedman M., Shen S., and Firon N., 2009, Transcriptional profiling of maturing tomato (Solanum lycopersicum L.) microspores reveals the involvement of heat shock proteins, ROS scavengers, hormones, and sugars in the heat stress response J. Exp. Bot., 60(13): 3891-3908 doi:10.1093/jxb/erp234
Fusco N., Micheletto L., Dal Corso G., Borgato L., and Furini A., 2005, Identification of cadmium-regulated genes by cDNA-AFLP in the heavy metal accumulator Brassica juncea L., J. Exp. Bot., 56(421): 3017-3027 doi:10.1093/jxb/eri299
Schaller G.E., and Anthony B., 1995a, Ethylene-binding sites generated in yeast expressing the arabidopsis ETR1 gene, Bleecker Science, 270: 1809
Schaller G.E., Andrea N.L., Michael B.L., Spanbauer J.M., and Anthony B.B., 1995b, The ethylene response mediator ETR1 from arabidopsis forms a disulfide-linked dimer, J. Biol. Chem., 270: 12526-12530 doi:10.1074/jbc.270.21.12526
Knee M., 1992, Sensitivity of ATPases to silver ions suggests that silver acts outside the plasma membrane to block ethylene action, Phytochemistry, 31:1093-1096 doi:10.1016/0031-9422(92)80240-F
Leymarie J., Bruneaux E., Gibot-Leclerc S., Corbineau F., 2007, Identification of transcripts potentially involved in barley seed germination and dormancy using cDNA-AFLP, J. Exp. Bot., 58(3):425-437 doi:10.1093/jxb/erl211
Liu Z.Y., Du Y.C., Wang X.X, Guo Y.M., and Gao J.C., 2008, Identification of differentially expressed genes by cDNA-AFLP approach during heat stress in tomato leaves, Yuanyi Xuebao (Acta Horticulturae Sinica), 35(7): 1011-1016 (刘志勇, 杜永臣, 王孝宣, 国艳梅, 高建昌, 2008, 高温胁迫下番茄叶片差异表达基因的cDNA-AFLP分析, 园艺学报, 35(7): 1011-1016)
Owens K.W., Peterson C.E., and Tolla G.E., 1980, Induction of perfect flowers on gynoecious muskmelon by silver nitrate and amino-ethoxyvinylglycine, HortScience, 15(5): 654-655
Qi Y.J., Cheng G.W., Wang C.S., and Wu L.J., 2004, The characteristics and chemical improvement of floral development of Cucurbita pepo L., Anhui Nongye Daxue Xuebao(Journal of Anhui Agricultural University), 31(4):452-455 (戚艺军, 程国旺, 王春生, 吴李君, 2004, 西葫芦花发育特性及其化学改良措施, 安徽农业大学学报, 31(4): 452-455)
Rodriguez F.I., Esch J.J., Hall A.E., Binder B.M., Schaller G.E., and Bleecker A.B., 1999, A copper cofactor for the ethylene receptor ETR1 from Arabidopsis, Science, 283: 996-998 doi:10.1126/science.283.5404.996
Snodgrass P.J., B.L. Vallee and Hockc F.L., 1960, Effects of silver and mercurials on yeast alcohol dehydrogenase, Journal of Biological Chemistry, 235: 504-508
Theresa F.L., Sabine L., and Sarah R.G., 2002, Silver enhances stamen development in female white campion (Silene latifolia), American Journal of Botany, 89 (6): 1014-1020 doi:10.3732/ajb.89.6.1014
Wang Q.M., and Zeng G.W., 1997, The effect of phytohormones and polyamines on sexual differentiation of momordica charantia, Yuanyi Xuebao (Acta Horticulturae Sinica), 24(1): 48-52 (汪俏梅, 曾广文, 1997, 激素和多胺对苦瓜性别分化的影响, 园艺学报, 24(1): 48-52)
Wang R.S., Li Y.R., Zhou S.M., Zhang M., Fang F.X., and Yang L.T., 2009, Stamen development and gene expression of flower of male-induced bitter melon gynoecious line, Xibao Shengwuxue Zazhi (Chinese Journal of Cell Biology), 31(4): 547-552 (王日升, 李杨瑞, 周生茂, 张曼, 方锋学, 杨丽涛, 2009, 纯雌系苦瓜花经AgNO3诱雄后的雄蕊发育和基因表达, 细胞生物学杂志, 31(4): 547-552)
Yang Y., Tang Y.Q., and Sakamoto M., 2004, Effect of chemical application and temperature on sex expression in bitter gourd (Monordica charantia L.), Redai Zuowu Xuebao (Chinese Journal of Tropical Crops), 25(3): 80-84 (杨, 唐燕琼, Sakaoto M., 2004, AgNO3,GA3和温度对苦瓜纯雌、雄株系性别分化的影响, 热带作物学报, 25(3): 80-84)